AATF Genome: Evolution of Allorecognition
Deepak Kaul
DOI10.21767/2573-5365.100010
Deepak Kaul*
Molecular Biology Unit, Experimental Medicine and Biotechnology Department, Postgraduate Institute of Medical Education and Research, Chandigarh-160012, India
- *Corresponding Author:
- Deepak Kaul
Molecular Biology Unit, Experimental Medicine and Biotechnology Department, Postgraduate Institute of Medical Education and Research, Chandigarh-160012, India
Tel: 91-172-755233
Fax: 91-172-2744401
E-mail: dkaul235@gmail.com
Received date: December 10, 2015; Accepted date: January 08, 2016; Published date: January 12, 2016
Citation: Kaul D. AATF Genome: Evolution of Allorecognition. Cell Mol Med. 2016, 2:1.
Abstract
Human AATF genome holds AATF gene and its encoded miR-2909 in its fold, to exhibit “Hysteresis phenomenon” arising out of bi-stable cooperative-activity of these AATF gene-products. This “Hysteresis Mechanism” enables cells, within human tissues, to decouple/couple the glycolysis from/with the aerobic respiration in a fashion that equips them with flexible dynamic programming to read the incoming signals as “Friend” or “Foe” and accordingly write the script for calibrated-response directed specifically/selectively to erase the signals perceived by these cells to be “Foe”. This novel model not only has the ability to explain the existing anomalies in “Allorecognition phenomenon” but may be even more useful for adding a new paradigm for futuristic studies directed to design preventive/ curative immunological-strategies against human diseases.
Keywords
Gene; Chromosome; Glucocorticoid
AATF Genome: Origin and Nature
Human AATF genome, of about 108 kb size, maps on chromosome 17 at 17q11.2-q12. Interestingly, in the same chromosomal region are several other genes encoding proteins that either have binding affinity for nucleic acids or act as chemokines including CCL5. AATF was, originally, identified as the translational-product of a transcript which was down-regulated by TGF-β in human intestinal cells [1] as well as interacting partner of human RNA polymerase II involved in cellular transcription [1]. AATF gene encodes phosphoprotein, containing 558 amino acids, that not only is highly conserved from yeast to man but is expressed in all the human tissues with stronger expression in brain, heart, thymus, kidney and placenta [1]. The amino acid sequence analysis of AATF protein revealed (Figure 1) the existence of a leucine zipper motif, two highly acidic amino-terminal domains, several phosphorylation sites and three nuclear reception binding LxxLL consensus sequences [1]. AATF biology assumed importance by the study that established a direct link between the ultrastructural features of AATF-mutant embryos and paucity of ribosomes, polyribosomes and rough endoplasmic reticulum [1] thereby raised the possibility that AATF protein may be involved in one or more aspects of the ontogenic control governing the embryonic development. The conspicuous presence of AATF protein within nucleus especially co-localized with nucleoli [1] reinforced the view that AATF protein may also function as a nucleolar stress sensor [1] for tailoring the immune response against a given sensed stress.

Figure 1: Characteristic features of AATF genome and its role in the cell decision.
A) AATF protein structure displaying domains containing Leucine Zippers, putative nuclear localization signals and
phosphorylation sites as well as having binding affinity for retinoblastoma gene product (RB1) and RNA Polymerase-II.
B) During G1-S cell cycle transition, AATF displaces HDAC1 from the Rb-E2F1 Complex to ensure activation of E2F-dependent promoters
and subsequent cell proliferation. AATF also activates p21 by displacing HDAC1 from the SP1 binding sites within p21 gene promoter.
Upon DNA damage, AATF is phosphorylated resulting in p53 expression and G2-M cell cycle arrest. AATF has the capacity to inhibit
apoptosis by interacting with several proteins (Dlk/Zip, Par-4 and NRAGE etc.).
C) The intronic-region, between exons 11 & 12 of AATF gene, is unique because this region not only codes for miR-2909 but also provides
many integration sites for HIV-1. Inhibition (denoted by 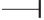 ). Stimulation (denoted by
). Stimulation (denoted by 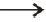 ).
).
AATF Interactome and Cell Decision
Cell Cycle regulators, having ability to influence both cell division and cell death, are involved in many physiological processes including tissue homeostasis, embryonic development and immune response [1]. AATF was discovered initially as an interacting molecular partner of subunit-11 of RNA polymerase-II as well as the retinoblastoma protein (Rb) which is the translational product of a first tumor suppressor gene cloned and identified from retinoblastoma tumors [1]. A major cellular target of Rb is the E2F family of transcription factors which regulate the genes involved in the cell-cycle progression for G1 to S-phase of the cellcycle [1,2]. The protein Rb was found to have binding affinity to both E2F and histone deacetylases (HDACs 1-3) and recruitment of HDAC-Rb-E2F Complex, at the promoters of genes involved in cellcycle transition G1-S phase, resulted in the arrest of cell growth at G1-phase of the cell-cycle [1]. AATF was found to have dual role in cell-cycle regulation because of its ability to displace HDAC1 not only from the Rb-E2F Complex resulting in E2F-dependent cell-cycle progression but also from SP1 transcription factor responsible for p21 expression thereby ensuring cell proliferation from G1 to S-phase as well as growth arrest at G1-phase of the cell-cycle [1]. Studies directed to resolve the involvement of AATF in the cellular response to the DNA damage revealed that AATF protein half-life was governed by the proteosomal pathway and stability by phosphorylation at four serine residues (Ser 187; Ser 141; Ser 474; and Ser 508) by the kinases ATM and ChK2 activated as a result of mammalian DNA damage [1]. Chromatin immuno-precipitation experiments revealed that in response to cellular DNA damage, AATF phosphorylation switches it from E2F target genes to increased occupancy at the p21 and p53 promoters [1], thereby ensuring the maintenance of cell-cycle G2/M check point essential to protect cells from DNA damage or oncogenic stress [1]. Importantly, AATF shares conserved motif (EExxxDDL) with several proteins involved in DNA damage response and repair leading to the general recognition that AATF may be critical component of oncogenic barriers. AATF protein, in phosphorylated form, gains affinity for NFkB family member RelA and AATF-RelA complex acquires the ability to induce p53 expression at the transcriptional level [1]. AATF Interactome also contains transcription co-activator acetylase p300 [3] which has been shown to acetylate PxxP motif in p53 leading to the transcription activation of p53 gene [4]. Acetylation of p53 on lysine residues, in the DNA binding and C-terminal regions, prevents its interaction with MDM2 leading to increase in the p53 protein stability [4]. Interestingly, IFN-γ induced p53 expression also employs the acetylase p300 in a fashion similar to AATF [5].
Further, binding affinity of AATF with p300 was also been found to enhance transactivation by several steroid hormone receptors including androgen, estrogen glucocorticoid receptors in a hormone-dependent manner [3]. Using STAT3 as an interacting partner, AATF was found to induce the transcriptional expression of Akt1 gene which, in turn, was responsible for regulating genes coding for KLF4, MDM2, C-myc, NFkB and IFN-γ [6-10]. Consequently, AATF has the potential to regulate several genes either through STAT3 or Akt 1. At this stage, it is pertinent to note that: a) IFN-γ has been shown to up-regulate the expression of genes coding for KLF4 and p53 [11,12] as well as down-regulate the c-myc gene transcription [13]; b) the tumor suppressor p53 has been found to up-regulate the expression of genes coding for KLF4 and MDM2 coupled with down-regulation of genes coding for C-myc, NFkB and IFN-γ [14-18]; c) the transcriptional factors E2F and C-myc were found induce the expression of genes coding for AATF [2,19] and polycomb group protein [20,21] Bmi-1 which, in turn, ensured sustained NFkB activation [22]; d) KLF4 was found to suppress SP1-dependent genes [19,21,23] coding for AATF, Bmi-1 and UCP2; e) C-myc has been shown to inhibit MDM2 protein through its ability to induce the expression of p19 ARF gene [24]. Taken together these findings precipitate the importance of AATF as the most crucial and critical cellcycle regulator as far was cell division is concerned. Importance of AATF in cellular apoptosis became apparent from the finding that revealed AATF-dependent inhibition of cellular apoptosis induced by DAP-like kinase (DLK), a member of the deathassociated protein (DAP) kinase family of pro-apoptosis serinethreonine kinases [1]. The anti-apoptotic role of AATF in neural tissues was revealed by its ability to bind Par4, a leucine Zipper protein that promotes neuronal cell death by down-regulation of Bcl-2 gene as well as aberrant production of β-amyloid peptide (Aβ) through the activation of β-secretase enzyme [25]. In human kidney proximal tubule cells, AATF over-expression resulted in the inhibition of apoptosis despite oxidative and apoptotic damage in renal tubular cells [26]. AATF was also shown to be a functional antagonist of cell-death inducer (NRAGE) during neuronal development, because its ectopic expression completely reverted NRAGE-induced cell death [1]. Recently, a new dimension was added to the belief that cell-cycle regulators are crucial for cell death by the finding which revealed that AATF had inherent capacity to put brakes on p53-driven apoptosis by binding to the PISMA, BAX and BAK promoters to repress the DNA damageinduced expression of these pro-apoptotic p53 target genes [27]. Strangely, AATF was found to be required for sustaining mutant p53 expression in several cancer cell lines and AATF depletion by siRNA was also accompanied by apoptosis in vitro and in vivo [1], thereby underlining the importance of AATF in survival of cells expressing mutant p53. AATF-biology assumed huge importance by the recent findings which revealed the conspicuous existence of a 23 KDa truncated AATF protein coupled with its unique Interactome, within human cancer cells derived from different tissue origin, responsible for sustained methylation of tumor suppressor genes thereby ensuring human cells to become and remain cancerous [28]. Thus, AATF has emerged to behave like a “Master Epigenetic Switch” that regulates cell cycle progression, check point control and apoptosis.
AATF RNome Hysteresis and Allorecognition Phenomenon
Circadian rhythm is a well-established physiological process that governs human behavioral traits, metabolism and body function. In this context, human AATF RNome assumes importance because it holds AATF coding-transcript and regulatory non-coding miR- 2909 within its fold to exhibit circadian rhythmicity [29] as a result of epigenetic cross-talk between AATF RNome and the mastergenes responsible for the regulation of host immunity, energy metabolism and cellular oncogenic/oncostatic activities (Figure 2). AATF protein ensures sustained expression of miR-2909 (Figure 2A) by the activation of NFkB through its ability to induce the expression of Akt gene directly as well as Bmi-I gene indirectly [1,20] which also influences p53 protein stability [30] and miR- 2909, on the other hand, ensures sustained expression of genes coding for Bmi-1, AATF, UCP2 and CCL5 [31,32]. However, AATF induced IFN-γ expression through Akt gene product [1,10] ensures repression of both the genes coding for AATF and miR-2909 (Figure 2A) thereby restoring the expression of genes coding for p53 and KLF4 as well as ensuring UCP2 gene down-regulation which, in turn, is accompanied by the activation of NFkB responsible for up-regulation of genes coding for AATF and miR-2909 (Figure 2A). Hence bi-stable expression of AATF gene and its encoded miR- 2909 ensures regulation of mitochondrial uncoupling protein (UCP2) in a cyclic fashion (Figures 2A and 2B). Several findings have suggested that UCP2 initiates aerobic-glycolysis by shunting pyruvate out of mitochondria through decoupling of glycolysis from aerobic-respiration [33]. Hence it becomes apparent that cyclic regulation of cellular energy metabolism from aerobicrespiration to aerobic-glycolysis would have been impossible to achieve without the existence of AATF RNome within human cells, which holds AATF mRNA and miR-2909 in a bi-stable expressional cooperative-state displaying a hysteresis characteristics (Figures 2 and 3). This hysteresis-phenomenon, exhibited by AATF RNome, has the inherent capacity to provide human cells with the ability to produce and secrete lactate in a periodic fashion (Figures 2 and 3). At this stage it is pertinent to note the findings that: a) Lactate has been shown to initiate nitric oxide (NO) production which, in turn, has been recognized to promote Th1 response coupled with IgA class-switching as well inhibit Th17 response [34-36]; b) UCP2 knock-out cells exhibit NFkB over-activation as well as increased “NO” production [37]; c) Nitric oxide was found to induce p53 gene expression [36]; d) p53 was found to downregulate eNOS transcription and induce iNOS transcriptional expression [38]; e) SP1 has the ability to induce CCL5 expression which plays an important role in p53 gene expression as well as IFN-γ production [39,40]; f) cellular CCL5 secretion is suppressed by PGE2 [41]; g) Exogenous “NO” causes over expression of TGF-β gene [42] which is known to inhibit IFN-γ expression [43]. Apart from periodic lactate production through AATF RNome-based hysteresis phenomenon, this mechanism also allows human cells to produce periodic fluxes of either INF-γ or IL-17 (Figures 2A and 2B). Interestingly, IFN-γ has the ability to induce the expression of KLF4 gene [11] which, in turn, regulates the differentiation of Th17 cells independently of RORγt [44]. IFN-γ has been shown not only to stimulate quiescent hematopoietic stem cells to proliferate and mobilize during chronic bacterial infection [45] but also involved in multiple aspects of iron metabolism and the immune response [46]. Further, most crucial function of nitric oxide in immune system has been widely reflected by its participation in protective immunity against various intracellular pathogens including viruses, bacteria and protozoa [47]. The cytotoxic activity of “NO’ has also been effective in host defense against tumor cells and alloantigens [48]. Hence AATF RNomedirected hysteresis phenomenon provides tissue-cells with flexible programming depending upon the time and space of their activation thereby ensuring functional plasticity of human cells to define continuously “Why”, “What” and “How” to respond to hostile pathogens, friendly gut microbes, nutritional components and heritable aberrations etc. This view is further strengthened by the HIV-1 induced CD4+ lymphopenia that is responsible for the morbidity and mortality caused by the opportunistic infections as well as neoplastic diseases [25] together with the ability of CCL5- dependent trans-membrane signaling to inhibit HIV-1 infection [49]. Interestingly, HIV-1 genome encoded microRNA designated as hiv1-miR-H1 was conspicuously able to suppress AATF gene expression by targeting its exon-1 [25]. However, the additional target of hiv1-miR-H1 in the intronic region between exon-11 and-12 of the AATF gene intrigued us to explore this region for the existence, if any, of a novel cellular miRNA that may have the ability to target HIV-1 genome. Such a study, indeed, revealed the existence of a novel miRNA (initially named as hmiR-Che-1 and now designated as miR-2909) that had the inherent capacity to target HIV-1 genome especially regions coding for hiv1-miR-H1 as well as Vpr gene [25]. Surprisingly, there are about ten HIV- 1 integration sites within the intronic-region spanning the DNAsequence that encodes miR-2909 (Figure 1) thereby revealing as to why it is critical for HIV-1 to suppress AATF gene expression together with to neutralize the cellular defense-armour in the form of miR-2909 encoded by the AATF genome [25] in order to cause CD4+ lymphopenia by employing CCR5 co-receptor for its entry into target cells [49]. Further, HIV-1 latency has posed major hurdle in devising various preventive or curative measures. Histone deacetylase (HDAC) inhibitors have, recently, shown promise in reactivating HIV-1 reservoirs [50]. In this context also, AATF gene assumes importance because AATF is known to displace HDAC and recruit co-activator acetylase p300 to various promoters in human genome [3]. Hence HIV-1 dependent suppression of AATF gene may contribute to its latency as well as pathogenesis of AIDS. On the basis of “AATF RNome Hysteresis Model”, three prominent aberrations can be postulated:-
Figure 2: Regulatory “Cross-talk” between AATF genome encoded AATF protein and miR-2909 involves the master-genes
responsible for the regulation of energy metabolism, host immunity and cellular oncogenic/oncostatic activities.
A) AATF protein ensures sustained expression of miR-2909 by the activation of NFkB through its ability to induce the expression of
Akt gene directly as well as Bmi-1 gene indirectly. Bmi-1, in turn, influences the p53 protein stability. On the other hand, miR-2909
ensures sustained expression of genes coding for Bmi-1, AATF, UCP2 and CCL5. However, AATF induced IFN-γ expression ensures
repression of both the genes coding for AATF and miR-2909 as well as ensuring UCP2 gene down-regulation which, in turn, is
accompanied by the activation of NFkB responsible for another surge in the expression of genes coding for AATF and miR-2909.
B) Up-regulation of UCP2 gene expression results in aerobic-glycolysis which, in turn, ensures sustain production of Lactate
responsible for nitric oxide production leading to predominant Th1 immune response accompanied by higher levels of TGF-β, IgA
class switching.
C) Activation of cellular KLF4 by the exogenous stimuli especially LPS, TNF-α, IFN-γ ensures sustained IL-17 expression and this
phenomenon is abolished by the presence of TGF-β.
D) Deregulated miR-2909 RNomics involving KLF4 leads to sustained levels of CYLD and IL-17 gene-products which ensures cellular
necroptosis to end in Autoimmune response.
E) The conspicuous existence of trunctated AATF protein within cancer cells ensures suppression of tumor suppressors p21 and
p53 through methylation of promoters. On the other hand, this truncated AATF protein ensures inhibition of p53-induced cellular
apoptosis and autophagy thereby forces the immune system to adopt the transformed cells/tumor through a mechanism prevailing
at the immune-privileged sites. Inhibition (denoted by or  ). Stimulation (donated by or
). Stimulation (donated by or 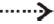 ).
).

Figure 3: The bistable expressional-relationship between AATF mRNA and miR-2909 within human cellular AATF RNome, can be best
described by the hysteresis loop: when AATF expression increases, the miR-2909 expression gradually increases to attain its
highest saturated level. As and when the AATF expression drops to zero-state, there still exists remnant miR-2909 expression.
However, when AATF expression falls to negative scale, the miR-2909 touches the lowest ebb. The rise of AATF expression again
is accompanied by the rise in miR-2909 expression thereby completing the hysteresis loop. This hysteresis–mechanism enables
cells, within human tissues, to decouple/couple the glycolysis from/with the aerobic-respiration in a fashion that equips these
cells with flexible programming to read the incoming signals and accordingly calibrate the immune-response for erasing the signals
perceived to be “Foe”. Expression level (High denoted by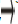 and low denoted by ).
and low denoted by ).
Sustained miR-2909 oscillations
Keeping in view the facts reported in Figures 2B and 2C, the transforming growth factor-β (TGF-β) has the inherent capacity to ensure sustained oscillations of miR-2909 through its ability to activate/deactivate NFkB in a cyclic fashion. The miR-2909 oscillations will, in turn, ensure the cellular production of lactate and CCL5 in a periodic fashion (Figure 1). Lactate-dependent “NO” production ensures the surge in TGF-β activity which, in turn, keeps in check the IFN-γ expression as well as secretion of CCL5 [42,51]. Generation of lactic acid also helps in the degradation of acid-labile IFN-γ which is a cytokine produced by natural killer (NK) cells of the innate immune system and by cytotoxic T lymphocytes [52] as well as one of the primary mediators of both innate and adaptive immunity [52]. Aerobic-glycolysis induced by miR-2909 can also disable the immune cells by using sugar [53]. Interestingly, immune privileged organs are well known to produce TGF-β as well as lactate [54-59] and it is their this ability which not only protects these organs from pathogens but also confers these organs with the inability to reject allogenic transplants even though these organs are bathed with the warmth and nutrients of blood (the mobile-tissue). Hence miR- 2909 oscillations ensure aerobic-glycolysis as the driving energysource for immune privileged organs to enable them to suppress Th1 type and DTH responses which is accompanied by increased recruitment of regulatory T-cells as well as induction of IgA classswitching.
Deregulated miR-2909 RNomics
This phenomenon depicted in Figure 2D can occur either by the mutation in the matured seed-sequence of miR-2909 or by mutation in the miR-2909 target 3’UTR region of the KLF4 mRNA resulting in the sustained expression of cellular KLF4 gene which, in turn, gives rise to sustained IL-17 production independently of RORγt [44] and CYLD gene expression [60]. IL-17 has the ability to induce cellular nitric oxide production which, in turn, can induce p53 gene expression [12,61] thereby again increasing KLF4 expression through p53 [14]. Interestingly, CYLD has been found to initiate programmed necrosis ”Necroptosis” which is non-apoptotic form of cell death that promotes the release of danger-associated molecular patterns (DAMPS) from necrotic cells to induce inflammation [62]. Recent compelling evidence showed that IL-17 producing cells are the major contributors to autoimmune disease [63]. Surge in IL-17 activity stimulates an enzyme called inducible nitric oxide synthesis (iNOS) to produce more nitric oxide that sets about destroying body tissue leading to autoimmune diseases [64]. Further, over-expressed KLF4 geneproduct down-regulates AATF expression which, not only has the ability to impair Th17 differentiation through Akt pathway [65] but also acts as a novel cyto-protective factor against oxidative cellular damage and death [26]. The salient features of the phenomenon (Figure 2)( are supported by the recent findings that reveal as to how human cells respond to high glucose or arsenic threat [31,66]. Further, deregulation of KLF4 expression also ensures restricted AATF-dependant control upon the p53- induced cellular apoptosis and autophagy [28,67]. Hence, cellular deregulated miR-2909 RNomics creates a situation that favours autoimmune response in tissue-specific/localized manner. This view is in conformity with the observed deregulated blood cellular miR-2909 RNomics in Rheumatoid Arthritis subjects [68].
Truncated AATF Protein
A new dimension was added by the recent finding that revealed the conspicuous existence of a 23 kDa truncated AATF protein within human cancer cells derived from different tissue origin [28]. This truncated AATF protein had the ability to recruit its unique interactome, consisting of partners such as SP1, DNMT3B and Par-4, to ensure repression of vital genes involved in tumor suppression as well as cell survival thereby defining the destiny of these cells to become and remain cancerous [28]. It can be stipulated from “AATF RNome Hysteresis Model” (Figures 2 and 3) that truncated AATF protein, observed in the cancer cells, can perform similar regulatory role to that performed by TGF-β in immune privileged sites and thereby can force the immune system to “adopt” the tumor as an immune-privileged novel organ. However, there is fundamental difference between TGF-β and truncated AATF protein as far as their role in the cell cycle regulation and cellular apoptosis is concerned. TGF-β inhibits primary tumor development and growth by inducing cell cycle arrest and apoptosis [69]. On the other hand, the truncated AATF protein promotes tumor development and growth by suppressing genes involved in tumor-suppression and activating genes that promote cellular immortality and growth [28]. Keeping in view the established fact that cancer cells exhibit DNA methylation addiction for achieving immortality and these cells consume glucose and produce lactic-acid under aerobic-Glycolysis condition [53], the role of truncated AATF protein assumes crucial importance because this protein ensures sustained aerobic-glycolysis in cancer cells to secrete lactate that not only insulates these cells from the cytotoxic NK Cells-and CD8+T-Cells but also starves the cytotoxic immune-cells to death through deprivation of glucose [53]. Hence ubiquitous presence of this truncated AATF protein within cancer cells can create a situation that favours immune privilege coupled with unbridled growth and immortality. This phenomenon, exhibited by cancer cells, explains as to why the immune system fails to reject tumors, even when many tumors do express new or mutated proteins [70]. Immunology has evolved for the last six decades or so, to believe that human immune system functions by discriminating “Self” (defined early in life) and “nonself” (imbibed later in life), sparing the “Self” and attacking the “non-self”. Keeping in view the “AATF RNome Hysteresis Model” proposed here (Figure 3), it becomes pertinent to ask as to how the immune privilege is extended to the antigens that emerged after puberty (spermatocyte antigens), during pregnancy (fetus antigens), or during lactation (milk antigens). The answer to this question evolves from the findings that revealed: a) Breast milk is a significant endogenous source of lactic acid-bacteria as well as contains TGF-β for initiation of IgA production in new born infants [58,59]; b) Lactate acts as a fetal fuel and TGF-β has been recognized to be of major importance in the scar-less wound repair observed in fetuses [56,57]; c) Spermatocytes utilize lactate as the main source of energy [54] and TGF-β signaling pathway is necessary to maintain spermatogenesis [55]. AATF RNome exhibited “Hysteresis Model” provides cells, within human tissues, with a unique mechanism to decouple/couple the glycolysis from/ with the aerobic-respiration in a fashion that allows these cells to undergo cyclic-dedifferentiation for expressing variety of receptors having capacity to read the incoming signals as “Friend” or “Foe” and write a script for calibrated-response in order to erase the signals perceived to be “Foe” by these cells. Further, cyclic cellular de-differentiation can also help the normal human cells to secrete exosomes enriched with newly changed tissueproteins (emerging as a result of puberty, metamorphosis and aging) as well as microRNAs and even MHC-peptide complexes. These exosomes, having phosphatidyl-serine (PS) on their outer membrane leaflet, can present an “eat-me” signal leading to “Apoptotic Mimicry”-a phenomenon exhibited by the cancer cells [71] to detract Nk Cells-and CD8+T cell-mediated cytotoxicity [71] as well as provide food for scavenging macrophages which also have the ability to secrete an immunosuppressive cocktail consisting of IL10, TGF-β and prostaglandin E2 [71]. Hence cyclic cellular de-differentiation, achieved by human cells during AATF RNome Hysteresis cycle (Figure 3), can explain as to how immune system achieves tolerance towards the antigens that appear after puberty or while aging or during metamorphosis or on oncogenic transformation.
Allorecognition: Concluding Remarks
All living organisms have evolved with the inbuilt survivalmechanism widely known as “Allorecognition phenomenon”. It is through this phenomenon, the organisms perceive “Faith” in their self (which includes friendly microbes and nutritional inputs to meet their energy requirements) and “Fear” from nonself (that includes hostile pathogens or metabolites or environmental factors having ability to pose stress/damage to self). Three models designated as self-nonself (SNS); Infectious-nonself (INS) and “Danger”, have been put forward so far to explain this “Allorecognition phenomenon” with limited/partial success in explaining various fundamental findings that define the functions of immune system. The “AATF RNome Hysteresis Model” proposed here has the inherent capacity not only to explain all the fundamental questions that have enveloped immunologists for the last six decades or so but also may be even more useful in defining new possibilities to understand the immune system in all its shades and dimensions. This “Hysteresis-Model” provides human tissues with flexible dynamic programming to sense and tailor their immune response to hostile pathogens, friendly gut microbes, nutritional components and heritable aberrations etc.
References
- Iezzi S, Fanciulli M (2015) Discovering Che-1/AATF: a new attractive target for cancer therapy. Front Genet 6:141.
- Mehrotra A, Joshi K, Kaul D (2010) E2F-1 RNomics is critical for reprogramming of cancer cells to quiescent state. Int J Cancer J Int Cancer 127: 849–858.
- Peter Leister SB (2003) Apoptosis antagonizing transcription factor AATF is a novel coactivator of nuclear hormone receptors. Signal Transduct 3:17 –25.
- Dornan D, Shimizu H, Burch L, Smith AJ, Hupp TR (2003) Theproline repeat domain of p53 binds directly to the transcriptional coactivator p300 and allosterically controls DNA-dependent acetylation of p53. Mol Cell Biol 23: 8846–8861.
- Contreras AU, Mebratu Y, Delgado M, Montano G, Hu C-AA, et al. (2013) Deacetylation of p53 induces autophagy by suppressing Bmf expression. J Cell Biol 201: 427–437.
- Sun Y, Zheng B, Zhang X, He M, GuoZ,et al. (2014) PPAR-γ agonist stabilizes KLF4 protein via activating Akt signaling and reducing KLF4 ubiquitination. BiochemBiophys Res Commun 443: 382–388.
- Ogawara Y, Kishishita S, Obata T, Isazawa Y, Suzuki T, et al. (2002) Akt enhances Mdm2-mediated ubiquitination and degradation of p53. J BiolChem 277: 21843–21850.
- Segrelles C, Moral M, Lara MF, Ruiz S, Santos M, et al. (2005)Molecular determinants of Akt-induced keratinocyte transformation. Oncogene 25: 1174–1185.
- Meng F, Liu L, Chin PC, DMello SR (2002) Akt is a downstream target of NF-kappa B. J BiolChem 277: 29674–29680.
- Okubo T, Washida K, Murakami A (2010) Phenethylisothiocyanate suppresses nitric oxide production via inhibition of phosphoinositide 3-kinase/Akt-induced IFN-gamma secretion in LPS-activated peritoneal macrophages. MolNutr Food Res 54: 1351–1360.
- Chen ZY, Shie JL, Tseng CC (2002) STAT1 is required for IFN-gamma-mediated gut-enriched Krüppel-like factor expression. Exp Cell Res 281: 19–27.
- Kano A, Watanabe Y, Takeda N, Aizawa TS (1997) Analysis of IFN-γ-Induced Cell Cycle Arrest and Cell Death in Hepatocytes. J Biochem 121: 677–683.
- Ramana CV, Grammatikakis N, Chernov M, Nguyen H, Goh KC, et al. (2000) Regulation of c-myc expression by IFN-gamma through Stat1-dependent and -independent pathways. EMBO J 19: 263–272.
- Yoon HS, Chen X, Yang VW (2003) Kruppel-like factor 4 mediates p53-dependent G1/S cell cycle arrest in response to DNA damage. J BiolChem278: 2101–2105.
- Barak Y, Juven T, Haffner R, Oren M (1993) mdm2 expression is induced by wild type p53 activity. EMBO J 12: 461–468.
- Ho JSL, Ma W, Mao DYL, Benchimol S (2005) p53-Dependent transcriptional repression of c-myc is required for G1 cell cycle arrest. Mol Cell Biol 25: 7423–7431.
- Lowe J, Shatz M, Resnick M, Menendez D (2013) Modulation of immune responses by the tumor suppressor p53. BioDiscovery.
- Chang CH, Curtis JD, Maggi LB, Faubert B, Villarino AV, et al. (2013) Posttranscriptional control of T cell effector function by aerobic glycolysis. Cell 153:1239–1251.
- Kaul D, Mehrotra A (2007) Functional characterization of AATF transcriptome in human leukemic cells. Mol Cell Biochem 297: 215–220.
- Nowak K, Kerl K, Fehr D, Kramps C, Gessner C, et al. (2006) BMI1 is a target gene of E2F-1 and is strongly expressed in primary neuroblastomas. Nucleic Acids Res 34:1745–1754.
- Wang H-B, Liu G-H, Zhang H, Xing S, Hu L-J, et al. (2013) Sp1 and c-Myc regulate transcription of BMI1 in nasopharyngeal carcinoma. FEBS J 280: 2929–2944.
- Jiang L, Song L, Wu J, Yang Y, Zhu X, et al. (2013) Bmi-1 promotes glioma angiogenesis by activating NF-κB signaling. PloS One 8: e55527.
- Jiang Y, Zhang H, Dong LY, Wang D, An W (2008) Increased hepatic UCP2 expression in rats with nonalcoholic steatohepatitis is associated with upregulation of Sp1 binding to its motif within the proximal promoter region. J Cell Biochem105: 277–289.
- Eischen CM, Weber JD, Roussel MF, Sherr CJ, Cleveland JL (1999) Disruption of the ARF-Mdm2-p53 tumor suppressor pathway in Myc-induced lymphomagenesis. Genes Dev 13:2658–2669.
- Kaul D, Ahlawat A, Gupta SD (2009) HIV-1 genome-encoded hiv1-mir-H1 impairs cellular responses to infection. Mol Cell Biochem 323:143–148.
- Xie J, Guo Q (2006) Apoptosis antagonizing transcription factor protects renal tubule cells against oxidative damage and apoptosis induced by ischemia-reperfusion. J Am SocNephrol JASN 17: 3336–3346.
- Höpker K, Hagmann H, Khurshid S, Chen S, Hasskamp P, et al. (2012) AATF/Che-1 acts as a phosphorylation-dependent molecular modulator to repress p53-driven apoptosis. EMBO J 31: 3961–3975.
- Sharma S, Kaul D, Arora M, Malik D (2015) Oncogenic nature of a novel mutant AATF and its interactome existing within human cancer cells. Cell BiolIntMar 39:326–333.
- Sharma S, Singh D, Kaul D (2015) AATF RNome has the potential to define post mortem interval. Forensic SciInt247:e21–24.
- Calao M, Sekyere EO, Cui HJ, Cheung BB, Thomas WD, et al. (2013) Direct effects of Bmi1 on p53 protein stability inactivates oncoprotein stress responses in embryonal cancer precursor cells at tumor initiation. Oncogene32:3616–3626.
- Kaul D, Sharma S (2015) High glucose-induced human cellular immune response is governed by miR-2909 RNomics. Blood Cells Mol Dis 54:342–347.
- Kaul D, Sharma S, Garg A (2015) Mitochondrial uncoupling protein (UCP2) gene expression is regulated by miR-2909. Blood Cells Mol Dis 55:89–93.
- Xu X, Duan S, Yi F, Ocampo A, Liu G-H (2013) Mitochondrial regulation in pluripotent stem cells. Cell Metab 18:325–332.
- Montoya JJ, Fernández N, Monge L, Diéguez G, Villalón ALG (2011) Nitric oxide-mediated relaxation to lactate of coronary circulation in the isolated perfused rat heart. J CardiovascPharmacol58:392–398.
- Cerutti A (2008) The regulation of IgA class switching. Nat Rev Immunol 8:421–434.
- Niedbala W, Cai B, Liu H, Pitman N, Chang L (2007) Nitric oxide induces CD4+CD25+ Foxp3 regulatory T cells from CD4+CD25 T cells via p53, IL-2, and OX40. ProcNatlAcadSci104:15478–15483.
- Niño Fong R, Fatehi-Hassanabad Z, Lee SC, Lu H, Wheeler MB, et al. (2011) Uncoupling protein-2 increases nitric oxide production and TNFAIP3 pathway activation in pancreatic islets. J MolEndocrinol46:193–204.
- Ying L, Hofseth LJ (2007) An emerging role for endothelial nitric oxide synthase in chronic inflammation and cancer. Cancer Res 67:1407–1410.
- Kumar D, Hosse J, von Toerne C, Noessner E, Nelson PJ (2009) JNK MAPK pathway regulates constitutive transcription of CCL5 by human NK cells through SP1. J ImmunolBaltimMd182:1011–1020.
- Sakthivel SK, Singh UP, Singh S, Taub DD, Igietseme JU, et al. (2008) CCL5 regulation of mucosal chlamydial immunity and infection. BMC Microbiol8:136.
- Qian X, Zhang J, Liu J (2011) Tumor-secreted PGE2 inhibits CCL5 production in activated macrophages through cAMP/PKA signaling pathway. J BiolChem286:2111–2120.
- Schmidt A, Geigenmueller S, Voelker W, Seiler P, Buddecke E (2003) Exogenous nitric oxide causes overexpression of TGF-beta1 and overproduction of extracellular matrix in human coronary smooth muscle cells. Cardiovasc Res 58:671–678.
- Lin JT, Martin SL, Xia L, Gorham JD (2005) TGF-beta 1 uses distinct mechanisms to inhibit IFN-gamma expression in CD4+ T cells at priming and at recall: differential involvement of Stat4 and T-bet. J ImmunolBaltimMd174:5950–5958.
- Lebson L, Gocke A, Rosenzweig J, Alder J, Civin C (2010) Cutting edge: The transcription factor Kruppel-like factor 4 regulates the differentiation of Th17 cells independently of RORγt. J ImmunolBaltimMd185:7161–7164.
- Baldridge MT, King KY, Boles NC, Weksberg DC, Goodell MA (2010) Quiescent haematopoietic stem cells are activated by IFN-gamma in response to chronic infection. Nature 465:793–797.
- Tomassini B, Arcuri G, Fortuni S, Sandi C, Ezzatizadeh V, et al. (2012) Interferon gamma upregulatesfrataxin and corrects the functional deficits in a Friedreich ataxia model. Hum Mol Genet 21:2855–2861.
- Pavanelli W, Silva J (2010)The Role of Nitric Oxide in Immune Response Against TrypanosomaCruzi Infection. Open Nitric Oxide 2:1–6.
- Wink DA, Vodovotz Y, Laval J, Laval F, Dewhirst MW, et al. (1998)The multifaceted roles of nitric oxide in cancer. Carcinogenesis 19:711–721.
- Tamamis P, Floudas CA (2014) Elucidating a key anti-HIV-1 and cancer-associated axis: the structure of CCL5 (Rantes) in complex with CCR5. Sci Rep4:5447.
- Smith PL, Tanner H, Dalgleish A (2014) Developments in HIV-1 immunotherapy and therapeutic vaccination. F1000prime Rep6:43.
- Huang WC, Yen FC, Shie FS, Pan CM, Shiao YJ, et al. (2010) TGF-beta1 blockade of microglial chemotaxis toward Abeta aggregates involves SMAD signaling and down-regulation of CCL5. J Neuroinflammation7:28.
- Schoenborn JR, Wilson CB (2007) Regulation of interferon-gamma during innate and adaptive immune responses. AdvImmunol96:41–101.
- Kim J, Dang CV (2006) Cancer’s molecular sweet tooth and the Warburg effect. Cancer Res 66:8927–8930.
- Jutte NH, Grootegoed JA, Rommerts FF, van der Molen HJ (1981) Exogenous lactate is essential for metabolic activities in isolated rat spermatocytes and spermatids. J ReprodFertil 62:399–405.
- Moreno SG, Attali M, Allemand I, Messiaen S, Fouchet P, et al. (2010)TGFbeta signaling in male germ cells regulates gonocyte quiescence and fertility in mice. DevBiol 342:74–84.
- Char VC, Creasy RK (1976) Lactate and pyruvate as fetal metabolic substrates. Pediatr Res 10:231–234.
- Cowin AJ, Holmes TM, Brosnan P, Ferguson MW (2001) Expression of TGF-beta and its receptors in murine fetal and adult dermal wounds. Eur J Dermatol EJD 11:424–431.
- Martín R, Langa S, Reviriego C, Jimínez E, Marín ML, et al. (2003) Human milk is a source of lactic acid bacteria for the infant gut. J Pediatr 143:754–758.
- Ogawa J, Sasahara A, Yoshida T, Sira MM, Futatani T, et al. (2004) Role of transforming growth factor-beta in breast milk for initiation of IgA production in newborn infants. Early Hum Dev 77:67–75.
- Kaul D, Arora M, Garg A, Sharma S (2015) MALT1 induced immune response is governed by miR-2909 RNomics. MolImmunol 64:210–217.
- Liu AC, Lee M, McManus BM, Choy JC (2012) Induction of endothelial nitric oxide synthase expression by IL-17 in human vascular endothelial cells: implications for vascular remodeling in transplant vasculopathy. J ImmunolBaltimMd188:1544–1550.
- Moquin DM, McQuade T, Chan FKM (2013) CYLD deubiquitinates RIP1 in the TNFα-induced necrosome to facilitate kinase activation and programmed necrosis. PloS One8:e76841.
- Zhu S, Qian Y (2012) IL-17/IL-17 receptor system in autoimmune disease: mechanisms and therapeutic potential. Clin Sci LondEngl122:487–511.
- Miljkovic D, Trajkovic V (2004) Inducible nitric oxide synthase activation by interleukin-17. Cytokine Growth Factor Rev 15:21–32.
- Bai D, Ueno L, Vogt PK (2009) Akt-mediated regulation of NFκB and the essentialness of NFκB for the oncogenicity of PI3K and Akt. Int J Cancer J Int Cancer 125:2863–2870.
- Kaul D, Sharma S, Sharma M, Arora M, Arora M (2014) Arsenic programmes cellular genomic-immunity through miR-2909 R Nomics. Gene 536:326–331.
- Malik D, Kaul D, Chauhan N, Marwaha RK(2014) miR-2909-mediated regulation of KLF4: a novel molecular mechanism for differentiating between B-cell and T-cell pediatric acute lymphoblastic leukemias. Mol Cancer13:175.
- Malik D, Sharma A, Raina A, Kaul D (2015) Deregulated Blood Cellular miR-2909 RNomics Observed in Rheumatoid Arthritis Subjects. Arch Med 7:1–6.
- Jakowlew SB (2006) Transforming growth factor-beta in cancer and metastasis. Cancer Metastasis Rev 25:435–457.
- Matzinger P, Kamala T (2011) Tissue-based class control: the other side of tolerance. Nat Rev Immunol 11:221–230.
- Berens C, Lauber K, Herrmann M (2013) Autoimmunity vs. cancer: predator vs. alien. Autoimmunity 46:287–293.

Open Access Journals
- Aquaculture & Veterinary Science
- Chemistry & Chemical Sciences
- Clinical Sciences
- Engineering
- General Science
- Genetics & Molecular Biology
- Health Care & Nursing
- Immunology & Microbiology
- Materials Science
- Mathematics & Physics
- Medical Sciences
- Neurology & Psychiatry
- Oncology & Cancer Science
- Pharmaceutical Sciences